
Конус нарастания, или точка роста стебля, занимает верхушечное положение. У наиболее примитивных листостебельных растений – мхов, хвощей – точка роста представлена лишь одной клеткой пирамидальной формы с округлым основанием, обращенным вверх. Все клетки, составляющие тело растения, — ее производные.
У цветковых и голосеменных растений точка роста имеет вид конуса с округлой вершиной, который состоит из множества меристематических клеток и спрятан в почке. Конус нарастания – самая активная часть побега. Здесь формируются первичная структура стебля, листья, боковые побеги, генеративные органы.
Конус нарастания состоит из верхушечной меристемы. Собственно верхушка стебля округла. Чуть ниже ее появляются небольшие выступы в форме серповидных валиков. Это зачатки листьев – примордии. Со временем валики увеличиваются в размерах, уплощаются и превращаются в настоящие листья. В пазухах этих листьев также рано образуются пазушные почки.
Детальное изучение конуса нарастания показало, что не все его участки равнозначны. По предложению А. Шмидта, в конусе нарастания различают тунику и корпус, по терминологии А.Имса, — мантию и сердцевину. Туника составляет наружные слои верхушечной меристемы, один или несколько. Деление клеток в ней идет перпендикулярно поверхности конуса, т.е. антиклинально, благодаря чему происходит рост верхушечной меристемы в поверхность. Корпус – центральная часть верхушечной меристемы. Клетки делятся во всех направлениях, что обусловливает объемный рост конуса нарастания. Граница между туникой и корпусом не всегда четкая. У голосеменных растений клетки, морфологически соответствующие тунике, делятся по всем направлениям.
Клетки корпуса неоднородны по толщине клеточной оболочки, по степени вакуолизации, по меристематической активности. По этим признакам можно выделить инициальные клетки (протомеристему), которые занимают верхушечное положение, и расположенные ниже стержневую и периферические зоны. В стержневой меристеме клетки вытянуты по оси, сильно вакуолизированы. Клетки периферической зоны мельче, форма неправильная, делятся во всех направлениях, меристематическая активность наибольшая. Стержневая и периферическая зоны верхушечной меристемы принимают участие в формировании первичной структуре стебля, органогенная роль (образование листьев, боковых побегов и репродуктивных органов) принадлежит периферической меристеме.
• Локальное отложение материалов клеточной стенки позволяет клеткам растений формировать длинные отростки
• В клетках, находящихся в точках роста, актиновые филаменты и микротрубочки обычно располагаются параллельно направлению роста побега
• Пучки актиновых филаментов направляют движение везикул к верхушке растения, где в процессе роста они сливаются с плазматической мембраной
• Количество и локализация клеток, способных образовывать точки роста, вероятно, находится под контролем микротрубочек
• Чтобы получить доступ внутрь растения симбиотические бактерии переключают на себя рост верхушки корневого волоска
Мы выяснили, что основные аспекты метаболической машины клеток растений посвящены синтезу компонентов клеточной стенки, которые постепенно добавляются к растущим боковым стенкам элонгирующих клеток. Такая форма диффузного роста представляет собой основной механизм формообразования растущих клеток растений, но не является единственной.
Некоторые специализированные клетки ограничивают рост небольшой площадью, позволяя формировать узкие отростки, у которых растет только верхушка. В таких точках роста клетки организованы совершенно по-другому, по сравнению с теми, которые участвуют в общей элонгации.
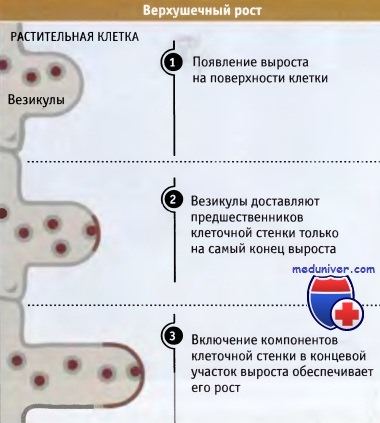 Верхушечный рост обеспечивается локальным характером доставки и слияния секреторных везикул,
Верхушечный рост обеспечивается локальным характером доставки и слияния секреторных везикул,
содержащих предшественники клеточной стенки, выделенные красным цветом.
Постоянный процесс слияния везикул с плазматической мембраной на конце выроста обеспечивает его рост,
вызывая удлинение бокового выроста при его неизменном диаметре.
Образование точек роста возможно благодаря локальной доставке предшественников клеточной стенки. Как показывает название, при удлинении отростка предшественники клеточных стенок и мембран доставляются только на самую его верхушку, имеющую форму конуса. В результате происходит удлинение клетки за счет удлинения ее боковых стенок, которое во много раз может превышать исходную длину.
Такой рост характерен для формирования корневых волосков и пыльцевых трубок. Как показано на рисунке ниже, развитие корневого волоска начинается с выпячивания на апикальном конце специализированной клетки эпидермиса (трихобласта), расположенной в зоне дифференцировки. Тонкие белые волоски, растущие перпендикулярно корню, существенно увеличивают площадь его поверхности, что способствует поглощению воды и ионов.
Образование пыльцевой трубки обеспечивает растительной клетке некую форму подвижности, которая необходима в некоторых критических условиях. У растений оплодотворение происходит после того, как пыльца от мужского цветка, с помощью насекомых или ветра, переносится на рыльце пестика женского цветка, который служит для нее как бы приемной антенной. Часто рыльце достигает больших размеров, что увеличивает вероятность получения пыльцы. Многие его части располагаются на некотором расстоянии от семязачатков цветка, содержащих яйцеклетки.
Имеющиеся на пыльцевых трубках точки роста позволяют им добраться до семязачатка. Как представлено на рисунке ниже, растущая пыльцевая трубка направляется вниз по рыльцу к семязачатку, куда она доставляет мужские гаметы, которые сливаются с яйцеклеткой, образуя диплоидный эмбрион. На рисунке ниже представлена частица пыльцы, которая образовала пыльцевую трубку.
Клетка в точке роста сильно поляризована. В то время как ее тело и большая часть отростка заняты вакуолями, сама точка роста и примыкающая к ней область заполнены цитоплазмой. В отростках присутствуют актиновые филаменты и микротрубочки, и, в отличие от общей элонгации клеток растений, они часто ориентированы параллельно направлению роста верхушки. На рисунке ниже показана ориентация актиновых филаментов в направлении роста верхушки.
В случае актиновых филаментов и микротрубочек, все филаменты имеют одинаковую полярность, и их быстрорастущие (плюс) концы располагаются ближе к кончику отростка. При такой организации цитоскелет обеспечивает доставку секреторных гранул на растущий кончик отростка. Молекулы миозина, связанные с поверхностью везикул, обеспечивают текучесть цитоплазмы, транспортируя везикулы в область, расположенную сразу же за точкой роста, где заканчиваются актиновые филаменты. Транспорт осуществляется вдоль стенок растущей трубки, и на конце везикулы сосредоточиваются в центре.
Этот процесс называется обратным фонтанирующим потоком. Как показано на рисунке ниже, на конце накапливается такая высокая концентрация везикул, которая исключает присутствие других органелл. Слияние везикул с плазматической мембраной обеспечивает дальнейший рост трубки. Поскольку актиновые филаменты играют основную роль в доставке везикул, они постоянно должны расти на конце трубки, чтобы дать ей возможность удлиняться.
Считают, что градиент ионов кальция, который образуется на конце пыльцевой трубки, регулирует связывание нескольких белков с актином. Это обеспечивает появление новых свободных концов филаментов, необходимых для полимеризации актина.
По сравнению с актином, роль микротрубочек в верхушечном росте менее понятна. Если в момент роста корневых волосков к ним добавить вещества, деполимеризующие микротрубочки, волоски продолжают расти зигзагообразно и в некоторых случаях даже формируют множественные точки роста. Поскольку любой рост трубочек свидетельствует о том, что доставка везикул продолжается, эти результаты позволяют предполагать, что микротрубочки как-то участвуют в пространственной координации верхушечного роста, однако не играют роли в процессах движения и слияния везикул.
Симбиотические азотфиксирующие бактерии проникают в овощные растения через растущие корневые волоски. Как показано на рисунке ниже, при прикреплении бактерии к корневому волоску он скручивается вокруг нее, напоминая пастушеский посох. Попав в такую ловушку, бактерия проникает в волосок. Рост его конца прекращается, и волосок поворачивается на себя в обратном направлении (напоминая вывернутый наизнанку палец на резиновой перчатке), образуя «инфекционную нить», которая тянется вдоль и проникает в клетку.
Для того чтобы вызвать такой искаженный рост, бактерии должна как-то реорганизовать процессы, связанные с доставкой везикул и их слиянием на конце волоска. При появлении инфекционной нити в теле клетки растения запускается цепь делений, в результате которых образуется клубенек, внутри которого поселяются колонии бактерий, снабжающие растение источниками связанного азота.
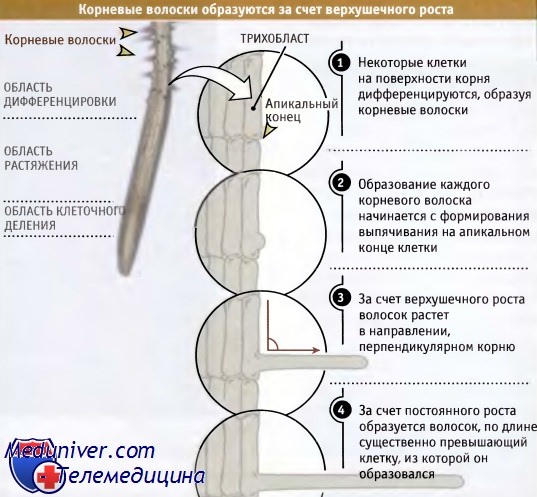 Слева представлен участок корня Arabidopsis.
Слева представлен участок корня Arabidopsis.
В процессе дифференцировки формируются корневые волоски, которые распространяются в стороны от поверхности корня.
Показано образование волоска. Каждый волосок представляет собой продукт вытягивания отдельной клетки. 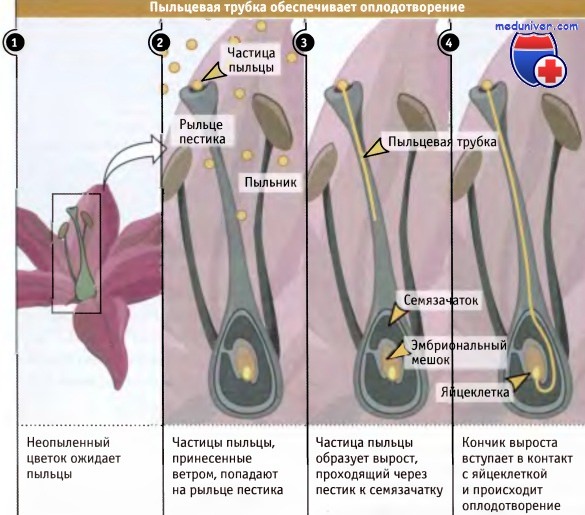 Верхушечный рост обеспечивает некоторым клеткам растений определенную подвижность.
Верхушечный рост обеспечивает некоторым клеткам растений определенную подвижность.
При опылении частицы пыльцы, захваченные цветком, образуют вырост за счет верхушечного роста достигающий яйцеклетки.
Длина выроста может во много раз превышать размер частицы пыльцы.  Частица пыльцы, образовавшая пыльцевую трубку.
Частица пыльцы, образовавшая пыльцевую трубку.
Сама частица представляет собой небольшую гранулу, которая на рисунке видна вверху слева.
Длина пыльцевой трубки может во много раз превосходить размеры самой пыльцевой частицы. 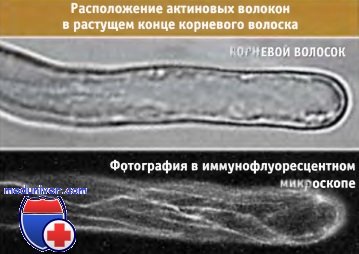 На верхней фотографии показан растущий конец корневого волоска.
На верхней фотографии показан растущий конец корневого волоска.
На нижней представлена та же область другого волоска,
актиновые волокна в котором содержат флуоресцентную метку.
Волокна ориентированы по длине. 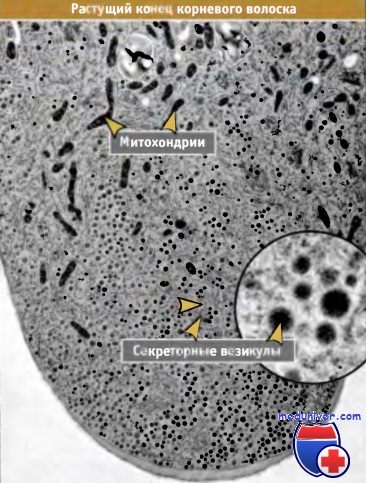 Фотография внутренней части растущего конца корневого волоска.
Фотография внутренней части растущего конца корневого волоска.
В этой части сконцентрированы секреторные везикулы, которые транспортируются с участием актинового цитоскелета.
Высокая концентрация везикул исключает присутствие в этой области других органелл, таких как митохондрии.
Везикулы постоянно сливаются с плазматической мембраной, обеспечивая рост. 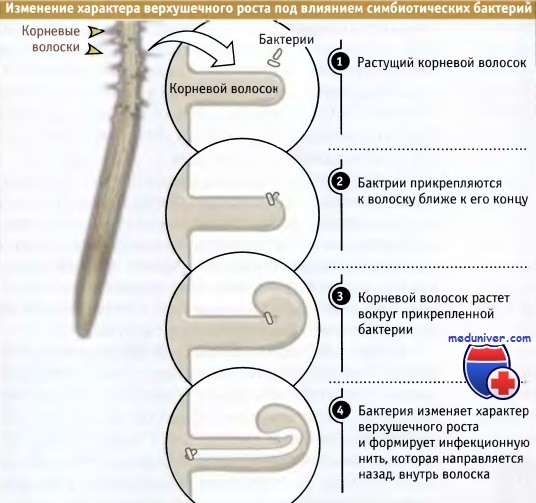 Последовательность событий, иллюстрирующая,
Последовательность событий, иллюстрирующая,
как симбиотические бактерии изменяют характер роста корневого волоска с тем,
чтобы проникнуть внутрь корня.
— В отличие от животных, имеющих (за немногими исключениями) ограниченный период Р., растения продолжают расти в течение всей своей жизни. Те из них, которые приносят плоды однажды в жизни (так называемые монокарпные растения) растут от семени до образования цветов, что может длиться различное время, смотря по растению: альпийские однолетники растут всего несколько недель, цветут и затем погибают; у так называемых столетников (виды Agave), принадлежащих также к монокарпным растениям, период Р. длится несколько десятков лет, но также прекращается с началом цветения. У многолетних растений, приносящих плоды неопределенное число раз, плодоношение не прекращает Р., последний неизменно продолжается из года в год, с временными остановками лишь в течение зимнего покоя; новые листья и новые молодые корни и ветви, с правильной периодичностью появляются на смену старым, и потому растения этой группы могут жить сотни и тысячи лет. Самая смерть их не является следствием глубокой старости, а обыкновенно происходит вследствие каких-либо случайных причин, как у знаменитого драконового дерева на острове Тенерифе, возраст которого определяли в 6000 лет и которое погибло (в 1868 г.) во время сильной бури. Эта особенность Р. растений удовлетворительно объясняется анатомическим строением их. В каком бы возрасте мы ни исследовали их, мы всегда найдем, кроме частей вполне выросших, еще такие, которые продолжают расти или даже находятся в зачаточном состоянии. Эти последние занимают строго определенные места — верхушки корня и стебля и их ветвей, как это представлено на приложенной схематической фигуре (фиг. 1), где наиболее затушеванными показаны те места, которые находятся в эмбриональном состоянии; это — так называемые точки возрастания растения; более слабой тушевкой отмечены части, которые уже вышли из эмбрионального состояния, но не достигли еще окончательного развития; наконец, части растения, окончившие свой Р., оставлены не затушеванными.

Фиг. 1. Схема распределения роста в двудольном растении: cc — семядоли; b, b’, b" — листья; k, k’, k" — почки; w, w’ — корни.
Строение точки возрастания в стебле и корне . На фиг. 2 точка возрастания стебля представлена в том виде, как она представляется под микроскопом. Она состоит из вполне однородных клеточек с тонкими оболочками, густо набитых плазмой и содержащих относительно крупные ядра.

Фиг. 2. Конус возрастания: A — хвоща; t — верхушечная клетка; S’, S" — сегментные клетки; f, f1 — зачатки листьев. B — Hippuris vulgaris; f — зачатки листьев; d — дерматоген; pr — периблема, pl — плерома. Остальное см. в тексте.
Клеточки эти находятся в постоянном делении, которое особенно наглядно выступает там, где есть особая так называемая верхушечная клеточка. Так бывает у споровых растений (мхов, хвощей, папоротников); на фиг. 3 изображен один из таких случаев. Верхушечная клетка имеет здесь форму трехсторонней пирамиды с выпуклым основанием, обращенным кверху. Она последовательно делится перегородками, параллельными трем сторонам, т. е. выросши до определенных размеров, она делится перегородкой параллельной одной стороне, затем снова вырастает до прежней величины и снова делится, но на этот раз перегородка образуется параллельно другой стороне и т. д.

Фиг. 3. Схема строения конуса возрастания, по Саксу.
Продукты деления ("сегментные клеточки"), вырастая, подвергаются дальнейшим делениям и дают начало всему конусу возрастания, состоящему из эмбриональной ткани, называемой первичной меристемой, верхушечная же клетка неизменно сохраняет свою величину и форму, являясь, таким образом, производительницей всех клеток тела растения; последнее является таким образом как бы продуктом деятельности этой удивительной клеточки. У высших, семенных растений особой верхушечной клетки не удалось найти, несмотря на многократные попытки; их конус (фиг. 2 A) возрастания состоит из вполне одинаковых меристематических клеточек, постоянно размножающихся делением. Происходящее отсюда, по-видимому, принципиальное различие в строении вегетативной верхушки споровых и семенных растений — по нашим основным представлениям, генетически связанных между собой, — сглаживается, однако, тем обстоятельством, что (как это впервые разъяснил Сакс) в обоих случаях процесс деления клеток подчинен собственно другому, более общему принципу — принципу взаимной перпендикулярности клеточных перегородок; каждая новая перегородка располагается так, чтобы пересекать прежде образовавшуюся под прямым углом. Принцип взаимной перпендикулярности перегородок входит в более общий — принцип наименьшей поверхности, согласно которому располагаются, например, и перегородки в мыльной пене. Осуществление последнего в растении важно уже в смысле экономии материала, так как клеточные оболочки строятся из такого ценного материала, как углеводы, которые растение должно само приготовлять для себя с большой затратой энергии. Присутствие или отсутствие верхушечной клетки представляет, следовательно, второстепенное значение: для растения важно лишь удовлетворять указанному выше принципу. Этот вывод можно подтвердить и фактами из сравнительной анатомии, которая показывает, что действительно, в пределах одного и того же рода (например, Lycopodium) и даже вида (L. Martensii) может быть верхушечная клетка и может не быть ее. Взаимная перпендикулярность клеточных перегородок, по Саксу, достигается в растении тем, что одни перегородки (переклинные) располагаются параллельно внешней поверхности органа, образуя своей совокупностью на продольном осевом разрезе конуса возрастания систему парабол (или линии иной кривизны), имеющих общую ось и общий фокус; другой род перегородок (антиклинные) на том же разрезе дает систему парабол, имеющих ту же ось и тот же фокус, но изогнутых в противоположном направлении. Наконец, третья система перегородок имеет радиальное направление. Если начертить на бумаге систему периклинных и антиклинных линий (см. фиг. 3), то получается схема, поразительно напоминающая продольный (медианный) разрез конуса возрастания (ср. фиг. 2 и 4).

Фиг. 4. Распределение роста в корне прорастающего гороха. Объяснение в тексте.
В некотором отдалении от верхушки взаимное давление клеток и залагающихся боковых органов вызывает различные смещения, вследствие чего указанная правильность в расположении перегородок нарушается. Строение конуса возрастания корня сходно с вышеописанным, с тем лишь различием, что в то время, как конус возрастания стебля бывает прикрыт молодыми листиками, в корне — лишенном листьев — имеется специальный покров для него — так называемый корневой чехлик. Закон деления клеток и здесь тот же. Подробнее см. Корень. Существуют и другие толкования строения конуса возрастания, например предложенное в 1865 г. Ганштейном. По этому взгляду, конус возрастания состоит из трех зародышевых пластов, так называемых гистогенов, надетых друг на друга наподобие колпачков. Наружный из этих гистогенов — дерматоген, состоящий из одного слоя клеток, впоследствии превращается в кроющую ткань растения (эпидерму); под дерматогеном находится несколько слоев клеток, образующих второй гистоген — периблему, превращающуюся впоследствии в кору стебля; наконец, третий гистоген, плерома, также состоит из нескольких слоев клеточек и занимает ось конуса возрастания; он дает начало главной массе стебля — сердцевине и проводящим пучкам. Действительно, иногда такое расчленение конуса возрастания выступает с большой ясностью, в большинстве же случаев тщательные исследования не могли открыть его.
Несколько ниже конуса возрастания начинается дифференциация постоянных тканей стебля. Прежде всего слой клеток, лежащих на поверхности, обособляется от остальных: клеточки его делятся исключительно перегородками, перпендикулярными к поверхности органа, и дают начало эпидерме. Одновременно с этим среди доселе однородной массы меристематической ткани появляются отдельные островки более мелких клеточек, сильно вытянутых в направлении продольной оси органа и образующих продольные шнуры; это так называемые прокамбиальные пучки, превращающиеся при дальнейшем развитии в проводящие пучки взрослого стебля. Вся остальная масса меристемы превращается частью в первичную кору, частью в сердцевину и сердцевинные лучи. О строении стебля и корня на этой стадии развития, равно как и о вторичном росте их в толщину — см. Стебель, Корень. Боковые органы — листья — залагаются очень рано, еще на самом конусе возрастания, в форме небольших выступов его (см. фиг. 2). Сколь ни малы эти зачатки (бугорки), в них намечена уже и будущая форма листа, т. е. будет ли он простой или сложный, цельнокрайный или раздробленный и т. д. Первоначально зачаток листа состоит из таких же меристематических клеток, как и конус возрастания стебля; при дальнейшем развитии в нем постепенно дифференцируются постоянные ткани, причем всего раньше Р. заканчивается в верхушке. Лист нарастает поэтому своим основанием, как это легко заметить на вытянутых в длину листьях многих однодольных. На конусе возрастания листовые зачатки залагаются очень близко друг к другу; молодые листики поэтому тесно смыкаются между собой и прикрывают голый конус возрастания, образуя так называемую верхушечную или главную почку. С возрастом, по мере развития междоузлий, листья раздвигаются. Если междоузлия развиваются слабо, получаются так называемые "розетки листьев", как у подорожников (Plantago). Очень рано, в углу, образуемом листовым зачатком с поверхностью главной оси, залагается новый конус возрастания, устроенный совершенно сходно с конусом возрастания главной оси. Это — угловая почка, зачаток боковой ветви стебля. Почка эта долго остается в зачаточном состоянии, между тем как лист продолжает развиваться. Поэтому боковые ветви появляются на более или менее значительном расстоянии от верхушки. Но если Р. главной оси почему-либо прекратится, например вследствие образования цветка или отмирания главной почки, то ближайшая угловая почка начинает усиленно развиваться, принимая характер главной. Таково происхождение симподиев, дихотомии (ложной) и пр. (см. эти слова).
Вышеописанный способ возрастания растений называется верхушечным Р.; но стебель может расти и в промежутках между верхушкой и основанием; такой способ возрастания и называется промежуточным или интеркалярным P. Общеизвестный пример в этом отношении представляют злаки, у которых в молодости листья бывают сильно сближены и своими влагалищами совершенно закрывают междоузлия; с возрастом последние сильно вытягиваются, так что листовые влагалища прикрывают теперь только нижнюю часть их; получается общеизвестная соломина. Это происходит оттого, что у этих растений, как и вообще у многих однодольных и некоторых двудольных, в основании каждого междоузлия существует особый пояс Р., устроенный аналогично верхушечному и закрытый снаружи листовым влагалищем. Благодаря деятельности меристематической ткани, здесь находящейся, каждое междоузлие самостоятельно вытягивается в длину нередко до весьма значительных размеров.
Физиология Р. Размножение клеток в конусе возрастания влечет за собой Р. органа (стебля или корня) в вертикальном направлении. Однако, в этой своей стадии, называемой стадией эмбрионального роста, удлинение органа еще незначительно. Если на корне прорастающего семени гороха нанести тушью черточки на расстоянии 1 мм друг от друга, то уже через 24 часа мы заметим, что, соответственно Р. органа, черточки сильно раздвинулись, но всего более те, которые находились не на самом конусе возрастания, а непосредственно за ним, а именно 2-ой и 3-ий мм Р., в смысле удлинения органа, происходит, следовательно, преимущественно в этой области. Объясняется это следующим образом. В конусе возрастания клетки имеют незначительную величину и густо набиты плазмой; они только размножаются делением. Несколько ниже с ними происходит замечательная перемена: в плазме их появляются полости, наполняющиеся водянистым соком — так называемые вакуоли. Вакуоли увеличиваются в объеме, сливаются друг с другом и, в то же время, жадно присасывая воду, возбуждают сильное осмотическое давление изнутри на клеточную оболочку. Последствием этого внутреннего давления (так называемого клеточного тургора) является растяжение оболочки и Р. ее в поверхности. Объем клетки сильно увеличивается, хотя количество собственно живого вещества (плазмы) остается, по-видимому, то же. Наружный вид клетки поэтому резко изменяется: размеры ее увеличиваются, протоплазма отходит к стенке и образует здесь тончайший постеночный мешочек, большая же часть полости заполняется огромной вакуолью, наполненной клеточным соком. Это — типический вид живой клетки растительных тканей. Вода является, следовательно, одним из важнейших строительных материалов растения, что составляет характерную черту их; дряблый вид завядших растений служит наглядным доказательством того, как много в прочности растения рассчитано на этот легко получаемый материал. Сущность процессов, происходящих в этой второй стадии роста, называется стадией растяжения, превосходно поясняется так называемыми искусственными клеточками Траубе. Эти искусственные клеточки могут быть сделаны из различных веществ, всего лучше из дубильно-кислого желатина. В раствор таннина погружают каплю желатина, предварительно подвергнутого кипячению до потери способности к застыванию. При соприкосновении желатина с таннином на поверхности капли тотчас осаждается коллоидальная перепонка из дубильно-кислого желатина; образуется, следовательно, замкнутый мешочек, внутри которого находится концентрированный раствор желатина; между этим раствором и раствором таннина устанавливается осмотический обмен через разделяющую их перепонку. Так как последняя очень плотна, то таннин не может пройти через нее, внутрь мешочка проходит только вода, и там накопляется значительное давление. Под этим давлением растягивается осадочная перепонка, и когда частицы ее достаточно разойдутся, желатин и таннин снова придут в соприкосновение и, соединившись, дадут новое количество осадка дубильно-кислого желатина. Таким образом, по мере растягивания искусственной клеточки внутренним давлением, в оболочку ее вставляются новые частицы, благодаря чему она и растет в поверхности. В общем, получается картина, поразительно напоминающая Р. растительных клеточек. Так как внутриклеточное давление (тургор), действительно, существует в живых растительных клеточках и может достигать значительной величины, то и принимают, что стадия растяжения клеточек в растении, подобно тому как и в опыте Траубе, является последствием двух механических причин: клеточного тургора и новообразования частиц клеточной оболочки. Под влиянием натяжения тургором, частицы клеточной стенки раздвигаются, и этим создаются условия для вставки новых частиц в промежутки между старыми. Такой способ Р. организованного образования известен под именем интусусцепции . В искусственных клеточках Траубе новые частицы перепонки образовались в момент встречи двух растворов, вследствие того, что растворенные в них вещества дают друг с другом нерастворимое соединение. Как и из чего в растительной клеточке образуются новые частицы оболочки — еще неизвестно. Стадия растяжения клеток сменяется стадией внутренней дифференциации. Потому ли, что клеточный тургор уменьшается, или потому, что к этому времени изменяются свойства клеточной оболочки (например, уменьшается ее растяжимость), но Р. оболочки в поверхности ослабевает, и новые частицы клеточки начинают отлагаться уже не в промежутках между старыми, а на них совнутри. Клеточная оболочка получает теперь разнообразную скульптуру, соответственно различным функциям растительного организма — происходит дифференциация постоянных тканей, чем и заканчивается Р. органа. Общая длина всех трех поясов Р. (эмбрионального, пояса растяжения и пояса внутренней дифференциации) в корне не превосходит 10 мм, в стебле значительно длиннее (несколько десятков см.). Быстрота Р. растений вообще довольно значительна, хотя конечно неодинакова у различных растений; особенно быстро растут листья Musa sapientium (1,1 мм в 1′). Много значат при этом и внешние условия — температура окружающей среды, свет и т. д. Начинаясь, вообще, при довольно низкой температуре, Р. растений усиливается с повышением ее, но только до известного предела, называемого оптимумом; при дальнейшем повышении температуры, Р. снова ослабевает и, наконец, совершенно останавливается, хотя растение остается еще живым и жизнедеятельным. Таким образом, влияние температуры на Р. определяется положением трех точек: minimum, optimum и maximum. В этом отношении различные виды растений сильно отличаются друг от друга, как видно из следующей таблицы:



